私は、近年話題となっているF1品種と固定種の話題に対して、比較的ニュートラルな立場をとっている。
もちろん、「固定種は自家採種できる」という面白さとか、「在来種の遺伝子自体に文化的価値がある」という意見に関しては、私も全面的にそう思うし、ソクラテスの煙草で販売しているタバコの種子はほとんどが固定種の種だ。
だが、「雄性不稔F1品種は子孫ができない不妊品種だから絶対ダメ。マジ危険」みたいな論調については、雄性不稔の仕組みをちゃんと理解したうえで、冷静に考えてみる必要があると思う。
母親の系統は花粉ができない。ならどうやって系統を維持するの?
少し考えてみれば、雄性不稔のF1品種が存在するなら、そして、雄性不稔が母株の細胞質(ミトコンドリア)を通して遺伝するなら、その母株はどうやって生まれたのか?母株の母株はどのように生まれたのか?という疑問が湧く。
この疑問に答えてくれる本が、私の本棚にあった。
『植物育種学 』鵜飼保雄著 東京大学出版 だ。
この本は、400ページ強の紙面すべてを育種の基本的な技術の解説にあてている、育種の入門書的な本だ。農学系の大学の授業などでよく使用されている。
本書の167ページに、わかりやすい図があったので、引用させていただく。
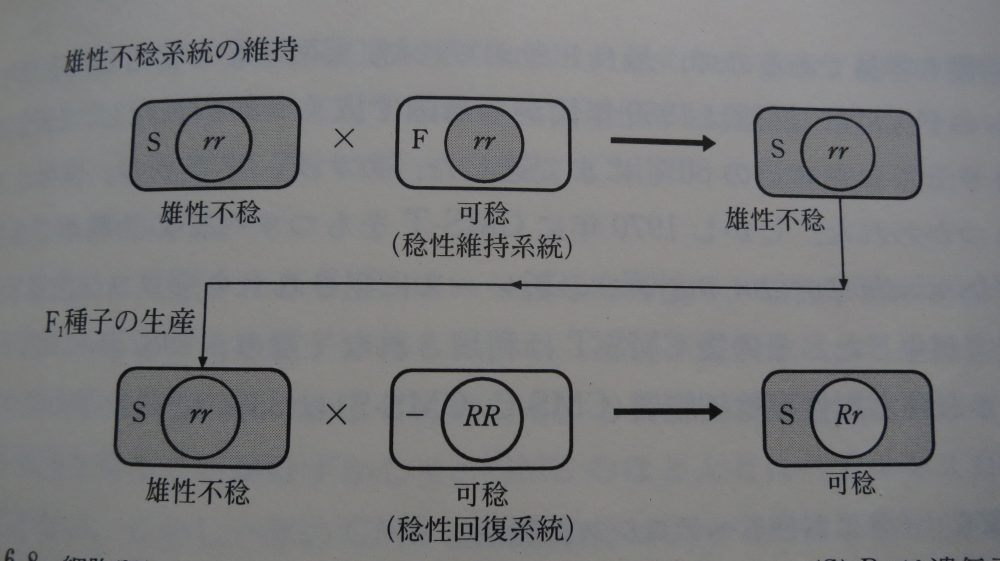
この図を見ただけで細胞質雄性不稔の仕組みがわかるのは生物マニアの方しかいないと思うので少し解説しよう。
まず、細胞質雄性不稔は細胞質遺伝子の異常だけでは発現しない現象である。
不稔細胞質(S)と、雄性不稔を引き起こす劣性の核遺伝子 rrの二つが存在して初めて、雄性不稔は目に見える形質として発現するのだ。
なので、たとえば不稔細胞質(S)-RRやRr の組み合わせ、または正常細胞質(F)-rrという組み合わせでは雄性不稔は発現せず、すべて可稔となる。
「雄性不稔F1品種の母親や、母親の母親はどうやって生まれたの?」という疑問は、この仕組みを理解することで解決できる。
雄性不稔F1品種の母親は、不稔細胞質(S)-rr だ。この母親の系統は花粉ができないから、純系系統として維持や大量増殖ができない。
そこで、稔性維持系統、つまり正常細胞質(F) を持つこと以外は母系統と同じかほぼ同じ可捻の系統「正常細胞質(F)-rr」を別に作っておき、その系統の花粉を不稔の母系統に掛け合わせることで、母系統を維持するのである。
母系統 (S)-rr はそのままでは花粉ができないが、隣に稔性維持系統(F)-rr を植えておけば、不稔の(S)-rr を維持することができる、というからくりだ。
雄性不稔のF1品種は子孫を残せる。でも、不稔が劣性形質だから・・・
上の図をみていただければわかるように、製品の段階、つまり我々が植えたり食べたりする個体はすべて画像右下の不稔細胞質(S)-Rrの組み合わせだから、花粉もできるし、種もできる。
なので「雄性不稔は種子ができない→危険」というのはそもそもの前提が間違っているし、少し煽り過ぎだと私は思う。
ただ、雄性不稔が自家採種の敵であることは間違いない。
そのことを説明するために、ここで、雄性不稔のF1品種から種を取る場合を想像してみよう。
まず、雄性不稔F1品種の個体の種(F2)を採ると、その種には、不稔細胞質-RR、不稔細胞質-Rr、不稔細胞質-rr が 1:2:1の割合で存在することになる。
これらのF2種子をたくさん蒔いて栽培すれば、可稔:不稔の割合は3:1になる。
不稔株は、「雄蕊が異常」という形質を目で確認できるから、rrの遺伝子型を持った株は人為的に抜き取ったりして、容易に除去できる。
rrの株を抜き取って除去したとしたら、畑に残った株の核の遺伝子型はRRとRrで、その比率は1:2となる。これら畑に残った株たちから再び種子を採ると(F3)、F3世代の種子にはRR:Rr:rrが、3:2:1の比率で存在することになる。
さらに、そのF3の種子を蒔いて栽培すると、可稔:不稔の割合は、5:1になる。
そして、これらのF3個体の中から、不稔株を抜き取って、また採種して・・・・
という風に繰り返していけば、不稔形質を持つ個体の出現割合は確かに低くなっていく。
だが、不稔形質が劣性遺伝子rによって引き起こされるがゆえに、不稔の核遺伝子rを集団から排除するのはなかなか面倒くさい。
なぜなら、Rrは見かけ上は可稔なので、「抜き取って排除する」ことができないからだ。
(ちなみに、正常細胞質ーrrの個体を個人が入手することは難しいから、検定交雑もできない。それをやる人がいるなら、ソイツはもうとっくに菜園家の域を超えてると思うw)
これは人間の病気でも同じで、劣性遺伝子による遺伝子疾患は、表現型としてあらわれにくいがゆえに、淘汰もされづらく、集団の中で地味に存在し続ける。
また、そもそも不稔細胞質というもの自体が正常ではないことは確かであり、「rrとの組み合わせで不稔になる」という因子を持った系統は、明らかに生存に不利である。
生命力というあいまいな言葉をあえて使うなら、雄性不稔F1品種は、「生命力の低い品種」ということができるだろう。
そんなわけで、自家採種にとって雄性不稔が面倒な存在となることは間違いないのである。
将来は固定種のほうが有利になるかも
現在のところ、モノの輸送にかかるエネルギーコストが安いから、F1品種で一斉にドカッと育てて、ドカッと消費地に運ぶほうが、地域で地産地消するより経済的だ。
だから、生育のばらつきが大きい固定種は農家にとって魅力がないし、自家採種するぐらいなら、その分の時間や労働をほかの作物の生産にあてたほうがいい。
しかし、将来、エネルギーコストが上がって、地産地消・自産自消のほうに分が上がるようになれば、少しずつ長期にわたって収穫できる固定種のほうが経済的になることもありうる。
そういう時代が来たときに、「ごめん、全部雄性不稔になっちゃってました。てへッ」となっていたら、それはなかなかまずいことだと思う。
われわれ菜園家は、コストをあまり気にしないでもいいという点で、ガチの農家たちよりは気楽な立場にいる。だからこそ、率先して、「種を採り、次世代に渡す」という責務を引き受けなければならないのかもしれない。
追記 2017/9/27
本稿の内容に一部補足があります。
「我々が植えたり食べたりする個体はすべて画像右下の不稔細胞質(S)-Rrの組み合わせだから、花粉もできるし、種もできる。」
という部分についてです。ここからちょいと長くなります。
さて、該当部分についてですが、あるF1個体の父親(記事内画像下段の真ん中)の細胞質遺伝子&核遺伝子の組み合わせが(F)-rrだった場合、そのF1個体は花粉を作ることができません。
これは盲点でした(反省)
イネやトウモロコシ、種子ができないと太らないトマトなどの作物以外の作物、つまり、葉を食べるキャベツやレタス、小松菜などは花粉ができなくとも問題はありませんので、父親の核遺伝子の遺伝子型がrrでも支障はありません。
しかしながら、父親の核遺伝子をrrにすることで何かメリットはあるのか?と考えると、わざわざ手間をかけてやることではない気もします。
雄性不稔はもともと「雑種強勢や斉一性を得たい→交雑が面倒→雄性不稔で半自動化できるやん!」という成り行きで開発された技術であり、そもそも、F1個体からF2世代の種子を採種することは考えてすらいないはずだからです。
さらに、自然界の原種や普通の品種たちの中にはrという劣性遺伝子はごくわずかしか存在しないのではないか?と私は思うのです。そうでなければ、最初の雄性不稔株はもっと簡単に見つかっていたはずだと考えられるからです。
少し単純化した例を挙げると、ほとんどのアフリカの人の皮膚の色は黒いですが、それは、彼らの遺伝子プールの中に「白い肌」という形質を作る劣性遺伝子がほとんど含まれないためです。
雄性不稔でも同じように、「不稔細胞質とセットで不稔になる」という形質をつくる劣性遺伝子rはごくまれにしか存在しないのではないでしょうか?
だとすると、父系統の遺伝子型をわざわざ珍しいrrに固定するっていうひと手間、要るのでしょうか?
耐病性や味覚など、ただでさえ調整しなければならない形質が大量にある中で、わざわざ「子孫を残せない」という形質を発現させる必要、あるかなあ?
まあ、「遺伝資源を奪われたくない」とか、「食料を支配して、ついでに人口削減してやるぜ!ギャハハハ!」という意地汚い理由または陰謀が狙いなら、それも考えられないことはないけど、不稔のF1株に他の可捻の株の花粉をかけて種子を採れば、遺伝資源を奪うことは全然可能なんですよね。
それに最近は、「雄性不稔F1は子孫が残せないらしい。なんかいかがわしいことやってるんじゃないか?」という話題が一部の菜園家の間で結構有名になりつつありますよね。種苗会社としては、要らぬ風評を避けるためにも、父親の核遺伝子をRRにして、そういう誤解を解いたほうが絶対お得だと思いませんか?
父親がRRなら、実際にF1個体(Rr)を育てて、花を咲かせて「ほら、花粉できるでしょ? 君たちが食べる段階の個体は、花粉も種子もできるよ。安心だよ。」と言えますからね。
そもそも、最近のF1品種vs固定種論争は、その発端に陰謀論者(陰謀論で人々を煽り、自分のとこでお金を使わせる人たち)が絡んでいたりいなかったりで、「F1は食料支配だ」という結論が独り歩きし過ぎてる感が否めない気がします。
でも実際のところ雄性不稔という技術は、
「雑種強勢を得るためにいろいろやってたら、雄性不稔が一番効率的だということが分かった!この機能マジ便利だわwいろんな品種に応用したろw」
ってな感じで進んでいった技術だと思うんですよね。
それに、自家採種が廃れたのも、めんどくさい&経済的じゃない、ってのが主な理由でしょう。誤解を恐れずに言えば、今の雄性不稔F1一強時代は我々菜園家の無知と怠慢が招いたことです。
F1が子孫を残せようが残せまいが、そもそもだれもF1個体から自家採種なんてしないんですよ。(え、私? 私はアイコというF1トマトからF2を採種して、数世代栽培し、ある程度固定した後、アントシアニン含有のトマトと掛け合わせて「俺のF1品種作出したったw」とかやってましたよ、ええ・・・)
まあそんなわけで、自家採種防止のためにわざわざコストをかけて父系統を(F)-rr にするメリットが種苗会社にはないのではないでしょうか?って私は思います。
はい、終わり!
ってことで、本稿の「雄性不稔は子孫残せる」という結論は一応そのまま堅持します。
だれか実験した人、又は、雄性不稔F1品種の父親の遺伝子型について詳しく知っている人がいたら、ぜひコメントお願いします。
タバコ種子はコチラ
8日に注文した2種類の種子は到着しています。迅速対応ありがとうございます。
出来れば採種に挑戦してみたいとも思いますが、異種類を交雑させない為にはどの程度離すべきでしょうか?
毎回質問してすみませんが、お時間のある時に回答頂けると幸いです。
今後ともよろしくお願いします。
質問にお答えいたします。
タバコ(ニコチアナ・タバカム)は、基本的に自分自身の花粉で自家受粉します。
が、昆虫が花粉を媒介することがあるので、交雑を完全に防ぐには、かなりの距離を離さないとだめだと思います。
数100m~とか、そういうレベルです。
しかしながら、不織布などでできた袋を花にかけることで、虫による交雑は防ぐことができます。
個人レベルの採種であれば数個の花を受粉させるだけで大丈夫なので、袋がけも簡単だと思います。
また何かありましたら、ブログでもメールでも、お気軽にお問い合わせください。